INAVA - INAVA




INAVA, иногда называемый гипотетический белок LOC55765, это белок неизвестной функции, которая у человека кодируется INAVA ген.[5] Менее распространенные псевдонимы генов включают FLJ10901 и MGC125608.
Ген
Место расположения

У человека INAVA располагается на длинной руке хромосома 1 в локус 1q32.1. Он охватывает от 200 891 499 до 200 915 736 (24,238 КБ) на положительной стороне.[5]
Джин соседство
INAVA фланкируется рецептором 25, связанным с G-белком (вверху) и членом семейства маэстро-теплоподобных повторов 3 (MROH3P), предполагаемым последующим псевдогеном. Рибосомный белок L34, псевдоген 6 (RPL34P6), находится дальше вверх по течению, а член семейства кинезинов 21B находится дальше вниз по течению.[5]
Промоутер

Существует семь предсказанных промоторов для INAVA, и экспериментальные данные свидетельствуют о том, что изоформа 1 и 2, наиболее распространенные изоформы, транскрибируются с использованием разных промоторов.[6] MatInspector, инструмент, доступный через Genomatix, использовался для прогнозирования фактор транскрипции сайты связывания в потенциальных промоторных областях. Факторы транскрипции, которые, как предполагается, нацелены на предполагаемый промотор изоформы 1, экспрессируются в ряде тканей. Наиболее распространенными тканями экспрессии являются мочеполовая система, нервная система и костный мозг. Это совпадает с данными по экспрессии белка INAVA, который высоко экспрессируется в почках и костном мозге.[7] Диаграмма прогнозируемой промоторной области с выделенными сайтами связывания транскрипционных факторов показана справа. Факторы, которые, как предполагается, связываются с промоторной областью изоформы 2, различаются, и двенадцать из двадцати прогнозируемых факторов экспрессируются в клетках крови и / или тканях сердечно-сосудистой системы.
Выражение
C1orf106 экспрессируется в широком диапазоне тканей. Данные экспрессии из профилей GEO показаны ниже. Сайты с наибольшей экспрессией перечислены в таблице. Экспрессия умеренная в плаценте, простате, яичках, легких, слюнных железах и дендритных клетках. Его мало в головном мозге, большинстве иммунных клеток, надпочечниках, матке, сердце и адипоцитах.[7] Данные по экспрессии из различных экспериментов, найденные на профилях GEO, предполагают, что экспрессия INAVA активируется при нескольких видах рака, включая рак легких, яичников, колоректального рака и груди.

| Ткань | Процентиль |
|---|---|
| В-лимфоциты | 90 |
| Трахеи | 89 |
| Кожа | 88 |
| Эпителиальные клетки бронхов человека | 88 |
| Колоректальная аденокарцинома | 87 |
| Почка | 87 |
| Язык | 85 |
| Поджелудочная железа | 84 |
| Приложение | 82 |
| Костный мозг | 80 |
мРНК
Изоформы
Девять предполагаемых изоформ продуцируются из гена INAVA, семь из которых, как предполагается, кодируют белки.[8] Изоформы 1 и 2, показанные ниже, являются наиболее распространенными изоформами.
Изоформа 1, самая длинная, принята в качестве канонической изоформы. Он содержит десять экзонов, которые кодируют белок длиной 677 аминокислот, в зависимости от источника. Некоторые источники сообщают, что белок состоит всего из 663 аминокислот из-за использования стартового кодона, который находится на 42 нуклеотида ниже по течению. Согласно NCBI, эта изоформа была предсказана только расчетным путем.[5] Это может быть потому, что Последовательность Козака окружение нижележащего стартового кодона больше похоже на консенсусную последовательность Козака, как показано в таблице ниже. Softberry использовали для получения последовательности предсказанной изоформы.[9] Изоформа 2 короче из-за усеченного N-конца. Обе изоформы имеют альтернативный сайт полиаденилирования.[8]

регуляция miRNA
miRNA-24 была идентифицирована как микроРНК которые потенциально могут нацеливаться на мРНК INAVA.[10] Сайт привязки, который находится в 5 'непереведенный регион Показано.
Протеин
Общие свойства

Изоформа 1, изображенная ниже, содержит домен DUF3338, две области низкой сложности и область, богатую пролином. Белок богат аргинином и пролином и содержит меньше, чем в среднем, аспарагина и гидрофобных аминокислот, особенно фенилаланина и изолейцина.[11] Изоэлектрическая точка составляет 9,58, а молекулярная масса немодифицированного белка составляет 72,9 кДал.[12] Предполагается, что белок не будет содержать N-концевой сигнальный пептид, но предполагается, что сигналы ядерной локализации (NLS) и богатый лейцином сигнал ядерного экспорта.[13][14][15]
Модификации
Предполагается, что INAVA будет сильно фосфорилироваться.[16][17] Сайты фосфоилирования, предсказанные PROSITE, показаны в таблице ниже. Прогнозы NETPhos показаны на диаграмме. Каждая линия указывает на предполагаемый сайт фосфорилирования и соединяется с буквой, которая представляет серин (S), треонин (T) или тирозин (Y).


Структура
Прогнозируется, что спиральные бухты будут охватывать остатки 130-160 и 200-260.[18] Предполагалось, что вторичный состав будет состоять примерно из 60% случайных витков, 30% альфа-спиралей и 10% бета-листов.[19]
Взаимодействия
Белки, с которыми взаимодействует белок INAVA, недостаточно хорошо изучены. Текстовый анализ данные свидетельствуют о том, что INAVA может взаимодействовать со следующими белками: DNAJC5G, SLC7A13, PIEZO2, MUC19.[20] Экспериментальные данные, полученные в результате двухгибридного скрининга дрожжей, предполагают, что белок INAVA взаимодействует с сигма-белком 14-3-3, который является адаптерным белком.[21]
Гомология
INAVA хорошо сохраняется у позвоночных, как показано в таблице ниже. Последовательности были получены из ВЗРЫВ[22] и BLAT.[23]
| Последовательность | Род и виды | Распространенное имя | Присоединение к NCBI | Длина (аа) | Идентичность последовательности | Время с момента расхождения (Mya) | |
|---|---|---|---|---|---|---|---|
| * | C1orf106 | Homo sapiens | Человек | NP_060735.3 | 667 | 100% | NA |
| * | C1orf106 | Macaca fascicularis | Макака-крабоядная | XP_005540414.1 | 703 | 97% | 29.0 |
| * | LOC289399 | Раттус норвегикус | Норвежская крыса | NP_001178750.1 | 667 | 86% | 92.3 |
| * | Прогнозируемый гомолог C1orf106 | Odobenus rosmarus divergens | Морж | XP_004392787.1 | 672 | 85% | 94.2 |
| * | C1orf106-подобный | Loxodonta africana | Слон | XP_003410255.1 | 663 | 84% | 98.7 |
| * | Прогнозируемый гомолог C1orf106 | Dasypus novemcinctus | Девятиполосный броненосец | XP_004478752.1 | 676 | 81% | 104.2 |
| * | Прогнозируемый гомолог C1orf106 | Ochotona princeps | Американская пищуха | XP_004578841.1 | 681 | 78% | 92.3 |
| * | Прогнозируемый гомолог C1orf106 | Monodelphis domestica | Серый короткохвостый опоссум | XP_001367913.2 | 578 | 76% | 162.2 |
| * | Прогнозируемый гомолог C1orf106 | Chrysemys picta bellii | Нарисованная черепаха | XP_005313167.1 | 602 | 56% | 296.0 |
| * | Прогнозируемый гомолог C1orf106 | Geospiza fortis | Средний наземный зяблик | XP_005426868.1 | 542 | 50% | 296.0 |
| * | Прогнозируемый гомолог C1orf106 | Аллигатор миссисипиенсис | Аллигатор | XP_006278041.1 | 547 | 49% | 296.0 |
| * | Прогнозируемый гомолог C1orf106 | Ficedula albicollis | Ошейниковая мухоловка | XP_005059352.1 | 542 | 49% | 296.0 |
| Прогнозируемый гомолог C1orf106 | Latimeria chalumnae | Латимерия Западной Индийского океана | XP_005988436.1 | 613 | 46% | 414.9 | |
| * | Прогнозируемый гомолог C1orf106 | Lepisosteus oculatus | Пятнистый гар | XP_006628420.1 | 637 | 44% | 400.1 |
| * | Домен FERM, содержащий 4A | Xenopus (Silurana) tropicalis | Западная когтистая лягушка | XP_002935289.2 | 695 | 43% | 371.2 |
| * | Прогнозируемый гомолог C1orf106 | Oreochromis niloticus | Нильская тилапия | XP_005478188.1 | 576 | 40% | 400.1 |
| Прогнозируемый гомолог C1orf106 | Haplochromis burtoni | Astatotilapia burtoni | XP_005914919.1 | 576 | 40% | 400.1 | |
| Прогнозируемый гомолог C1orf106 | Пундамилия Ньеререй | Haplochromis nyererei | XP_005732720.1 | 577 | 40% | 400.1 | |
| * | LOC563192 | Данио Рерио | Данио | NP_001073474.1 | 612 | 37% | 400.1 |
| LOC101161145 | Oryzias latipes | Японская рисовая рыба | XP_004069287.1 | 612 | 33% | 400.1 |
График идентичности последовательности в зависимости от времени, прошедшего с момента расхождения для записей, отмеченных звездочкой, показан ниже. Цвета соответствуют степени родства (зеленый = близкое родство, фиолетовый = дальнее родство).

Паралоги
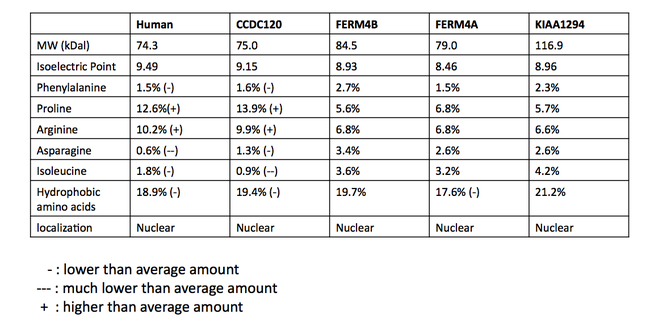
Белки, которые считаются паралогами INAVA, не согласованы между базами данных. Множественное выравнивание последовательностей (MSA) потенциально паралоговых белков было сделано, чтобы определить вероятность истинно паралогичных отношений.[24] Последовательности были получены в результате поиска BLAST у людей с белком C1orf106. MSA предполагает, что белки имеют общий гомологичный домен, DUF3338, который обнаружен у эукариот. Часть выравнивания множественных последовательностей показана ниже. Не считая домена DUF (выделено зеленым), сохранения было мало. Домен DUF3338 не обладает какими-либо экстраординарными физическими свойствами, однако одним примечательным открытием является то, что каждый из белков в MSA, по прогнозам, имеет два сигнала ядерной локализации. Предполагается, что все белки в MSA локализуются в ядре.[13] Сравнение физических свойств белков также было проведено с использованием SAPS и показано в таблице.[11]

Клиническое значение
Всего 556 однонуклеотидный полиморфизм (SNP) были идентифицированы в области гена INAVA, 96 из которых связаны с клиническим источником.[25] Rivas et al.[26] идентифицировали четыре SNP, показанные в таблице ниже, которые могут быть связаны с воспалительное заболевание кишечника и болезнь Крона. Согласно GeneCards, ассоциации с другими заболеваниями могут включать: рассеянный склероз и язвенный колит.[27]
| Остаток | Изменять | Примечания |
|---|---|---|
| 333 (rs41313912) | Тирозин ⇒ фенилаланин | Фосфорилированный, умеренная консервация |
| 376 | Аргинин ⇒ цистеин | Умеренное сохранение |
| 397 | Аргинин ⇒ треонин | Не сохранилось |
| 554 (RS61745433) | Аргинин ⇒ цистеин | Умеренное сохранение |
Модельные организмы
Модельные организмы были использованы при изучении функции INAVA. Условный нокаутирующая мышь линия называется 5730559C18Riktm2a (EUCOMM) Wtsi был создан на Wellcome Trust Sanger Institute.[28] Самцы и самки животных прошли стандартизованный фенотипический скрининг[29] для определения последствий удаления.[30][31][32][33] Проведены дополнительные проверки: - Углубленное иммунологическое фенотипирование[34] - углубленное фенотипирование костей и хрящей[35]
| Характеристика | Фенотип |
|---|---|
| Все данные доступны на сайте.[29][34][35] | |
| Лейкоциты периферической крови 6 недель | Нормальный |
| Инсулин | Нормальный |
| Гематология 6 недель | Нормальный |
| Гомозиготная жизнеспособность на P14 | Нормальный |
| Гомозиготная фертильность | Нормальный |
| Масса тела | Нормальный |
| Неврологический осмотр | Нормальный |
| Сила захвата | Нормальный |
| Дисморфология | Нормальный |
| Косвенная калориметрия | Нормальный |
| Тест толерантности к глюкозе | Нормальный |
| Слуховой ответ ствола мозга | Нормальный |
| DEXA | Нормальный |
| Рентгенография | Нормальный |
| Морфология глаза | Нормальный |
| Клиническая химия | Нормальный |
| Гематология 16 недель | Нормальный |
| Лейкоциты периферической крови 16 недель | Нормальный |
| Вес сердца | Нормальный |
| Сальмонелла инфекционное заболевание | Нормальный |
| Цитотоксическая функция Т-лимфоцитов | Нормальный |
| Иммунофенотипирование селезенки | Нормальный |
| Иммунофенотипирование мезентериальных лимфатических узлов | Нормальный |
| Иммунофенотипирование костного мозга | Нормальный |
| Эпидермальный иммунный состав | Нормальный |
| Проблема гриппа | Нормальный |
Рекомендации
- ^ а б c ГРЧ38: Ансамбль выпуск 89: ENSG00000163362 - Ансамбль, Май 2017
- ^ а б c GRCm38: выпуск Ensembl 89: ENSMUSG00000041605 - Ансамбль, Май 2017
- ^ "Справочник человека по PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ "Ссылка на Mouse PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ а б c d "NCBI Gene 55765". Получено 10 февраля 2014.
- ^ «Геноматикс: МатИнспектор». Получено 6 марта 2014.
- ^ а б «Профили GEO». Получено 6 марта 2014.
- ^ а б "Aceview". Получено 6 марта 2014.
- ^ «Мягкая ягода». Получено 20 апреля 2014.
- ^ "TargetScanHuman 6.2". Получено 15 апреля 2014.
- ^ а б «Статистический анализ белковых последовательностей». Получено 20 апреля 2014.
- ^ «Инструмент вычисления pI / Mw». Получено 10 апреля 2014.
- ^ а б «ПСОРТII». Получено 20 апреля 2014.
- ^ "cNLS Mapper". Получено 20 апреля 2014.
- ^ «НетНЭС». Получено 20 апреля 2014.
- ^ «НЕТФОС». Получено 20 апреля 2014.
- ^ «Швейцарский институт биоинформатики: PROSITE».
- ^ «КАТУШКИ EXPASY». Получено 20 апреля 2014.
- ^ «СОПМА». Получено 27 апреля 2014.
- ^ "НИТЬ". Получено 15 апреля 2014.
- ^ «МЯТА». Получено 15 апреля 2014.
- ^ "ВЗРЫВ". Получено 8 марта 2014.
- ^ «БЛАТ». Получено 8 марта 2014.
- ^ "Инструментальные средства биологии SDSC: ClustalW". Получено 12 марта 2014.
- ^ "dbSNP". Получено 22 апреля 2014.
- ^ Ривас М.А.; и другие. (2011). «Глубокое ресеквенирование локусов GWAS позволяет выявить независимые редкие варианты, связанные с воспалительным заболеванием кишечника». Природа Генетика. 43 (11): 1066–1073. Дои:10,1038 / нг.952. ЧВК 3378381. PMID 21983784.
- ^ «Генные карты». Получено 1 мая 2014.
- ^ Гердин А.К. (2010). «Программа генетики мыши Сэнгера: характеристика мышей с высокой пропускной способностью». Acta Ophthalmologica. 88: 925–7. Дои:10.1111 / j.1755-3768.2010.4142.x.
- ^ а б «Международный консорциум по фенотипированию мышей».
- ^ Скарнес В.К., Розен Б., Вест А.П., Кутсуракис М., Бушелл В., Айер В., Мухика А.О., Томас М., Харроу Дж., Кокс Т., Джексон Д., Северин Дж., Биггс П., Фу Дж., Нефедов М., де Йонг П.Дж., Стюарт AF, Брэдли А. (июнь 2011 г.). «Ресурс условного нокаута для полногеномного исследования функции генов мыши». Природа. 474 (7351): 337–42. Дои:10.1038 / природа10163. ЧВК 3572410. PMID 21677750.
- ^ Долгин Э (июнь 2011 г.). "Библиотека мыши настроена на нокаут". Природа. 474 (7351): 262–3. Дои:10.1038 / 474262a. PMID 21677718.
- ^ Коллинз Ф.С., Россант Дж., Вурст В. (январь 2007 г.). «Мышь по всем причинам». Клетка. 128 (1): 9–13. Дои:10.1016 / j.cell.2006.12.018. PMID 17218247.
- ^ White JK, Gerdin AK, Karp NA, Ryder E, Buljan M, Bussell JN, Salisbury J, Clare S, Ingham NJ, Podrini C, Houghton R, Estabel J, Bottomley JR, Melvin DG, Sunter D, Adams NC, Sanger Institute Проект генетики мышей, Таннахилл Д., Логан Д.В., Макартур Д.Г., Флинт Дж., Махаджан В.Б., Цанг С.Х., Смит I, Ватт FM, Скарнес В.К., Дуган Джи, Адамс DJ, Рамирес-Солис Р., Брэдли А., Сталь КП (2013) . «Полногеномное поколение и систематическое фенотипирование мышей с нокаутом открывает новые роли для многих генов». Клетка. 154 (2): 452–64. Дои:10.1016 / j.cell.2013.06.022. ЧВК 3717207. PMID 23870131.
- ^ а б «Консорциум иммунофенотипирования инфекций и иммунитета (3i)».
- ^ а б «Консорциум OBCD».
внешняя ссылка
- Человек C1orf106 расположение генома и C1orf106 страница сведений о генах в Браузер генома UCSC.